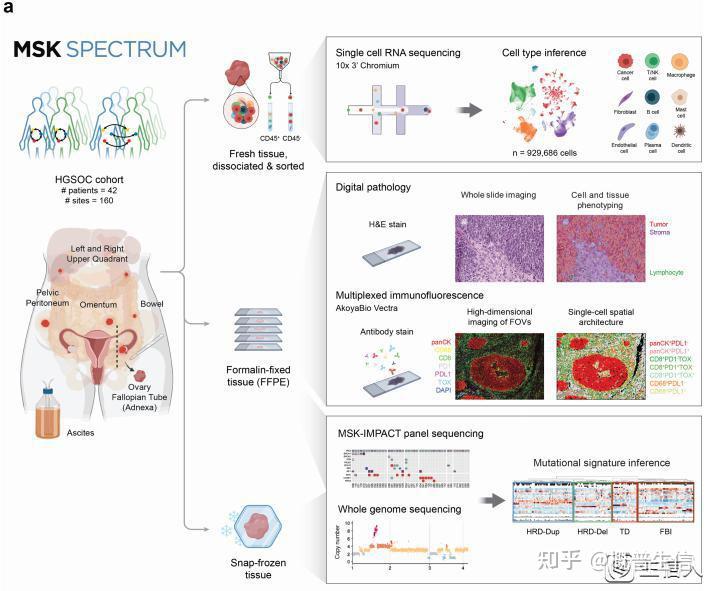
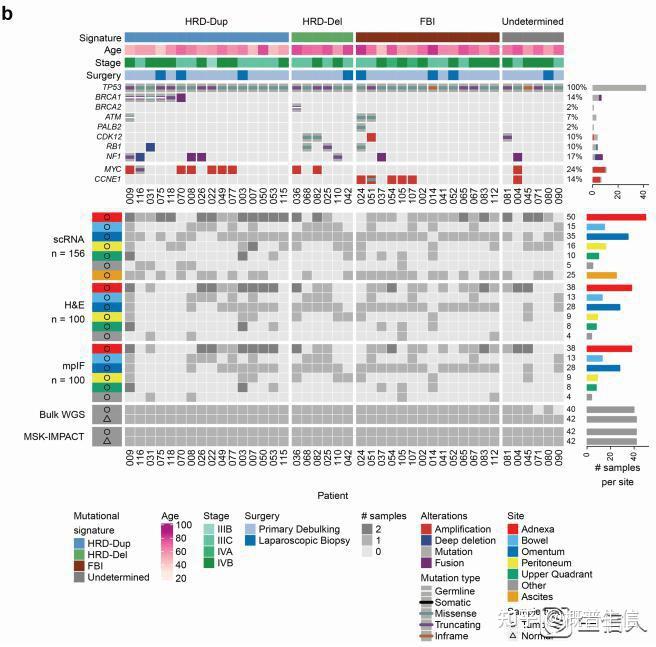
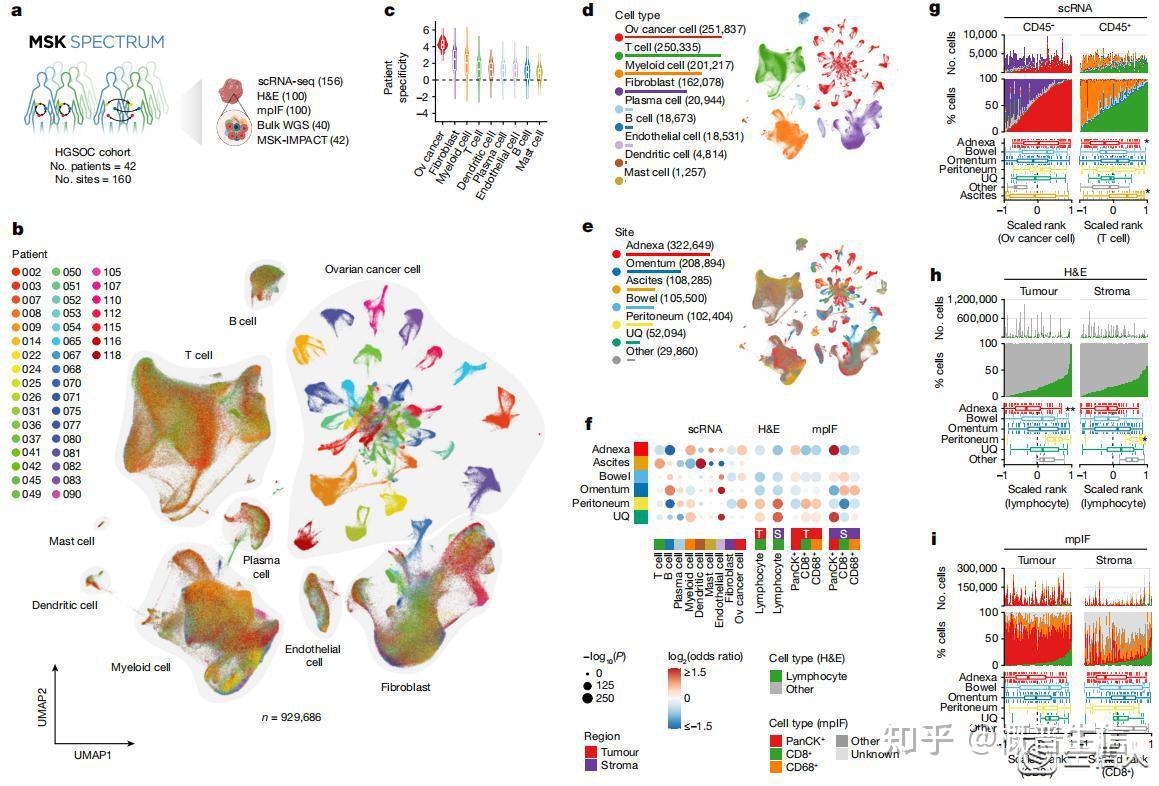
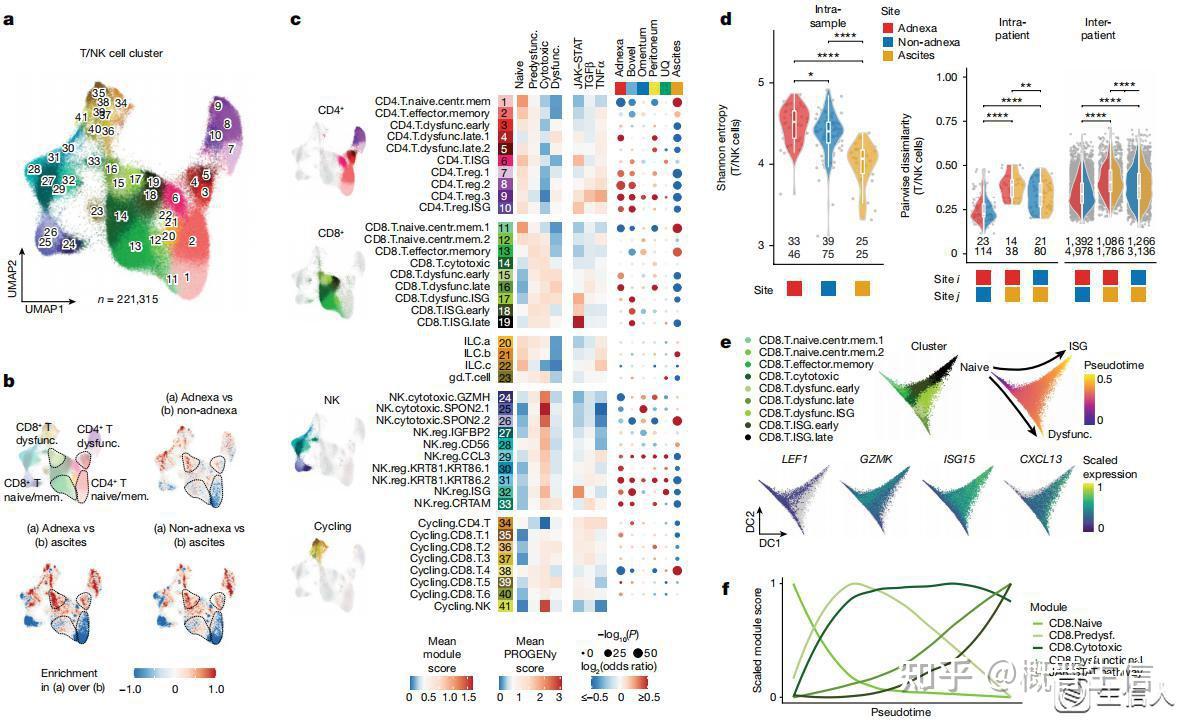
2
6
新手上路
使用道具 举报
0
9
18
7
14
本版积分规则 发表回复 回帖后跳转到最后一页
Archiver|手机版|小黑屋|同城医药问答网
GMT+8, 2025-12-24 19:22 , Processed in 0.474853 second(s), 21 queries .
Powered by Discuz! X3.4
© 2001-2013 Comsenz Inc.



 发表于 2023-1-12 08:51:41
发表于 2023-1-12 08:51:41